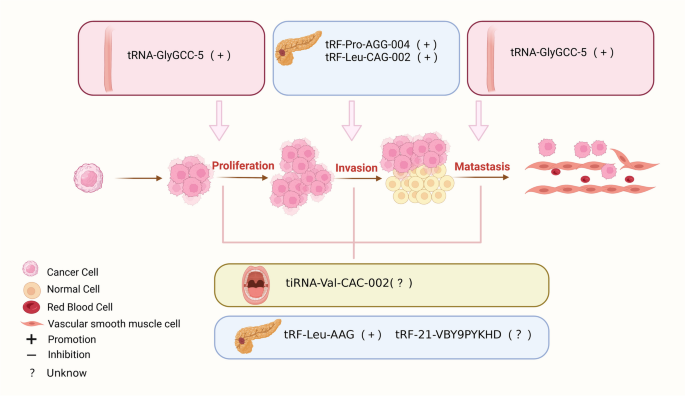
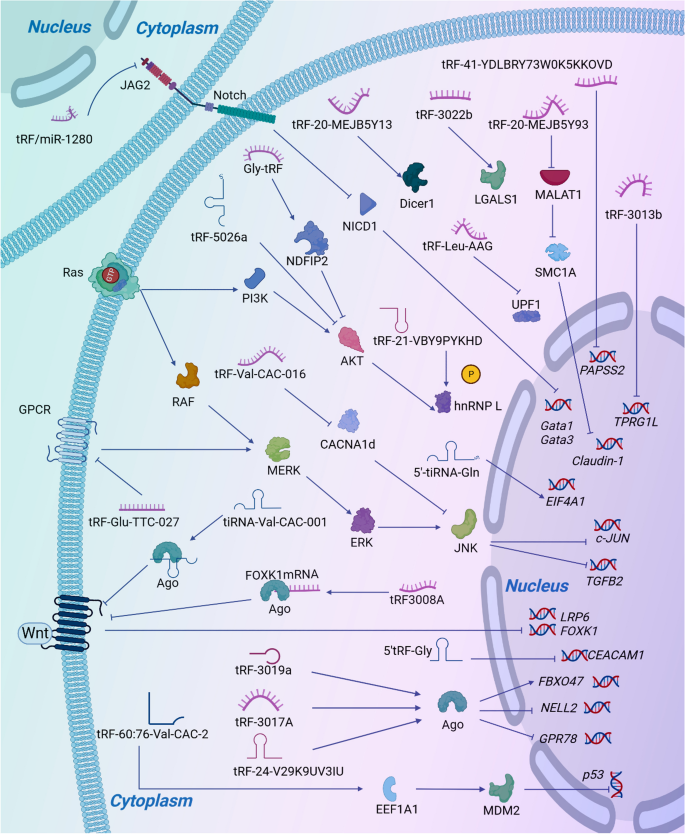
tsRNAs是源自tRNA的小型非编码RNA,在RNA沉默、核糖体生物发生、反转录转座、表观遗传等多种生理活动中发挥重要作用,并参与细胞分化、增殖和凋亡。tsrna相关异常通过影响细胞周期和特异性信号分子,对包括恶性肿瘤在内的许多人类疾病的发生、发展和进展具有重要影响。本文介绍了tsRNAs的起源和分类,综述了tsRNAs在消化系统疾病中的调节机制、生理功能和功能失调作用,并重点介绍了tsRNAs作为诊断和预后生物标志物的临床前景。
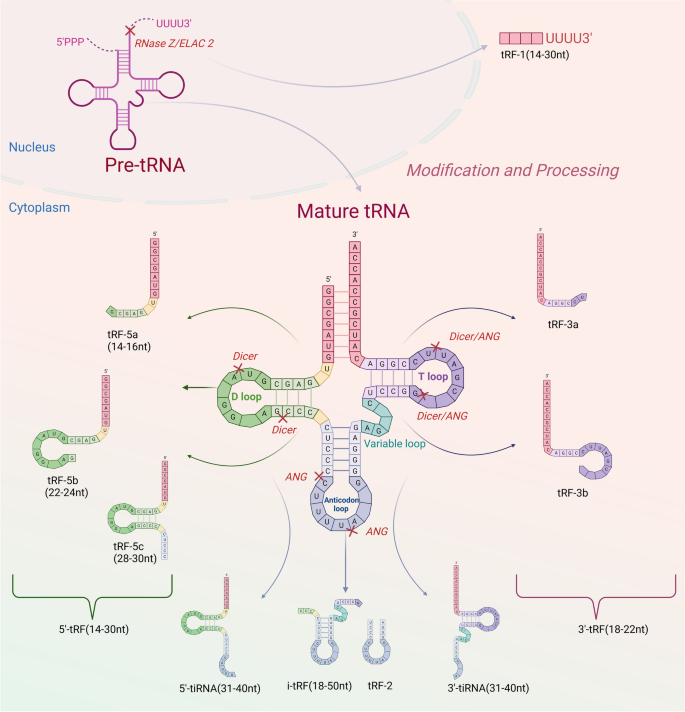
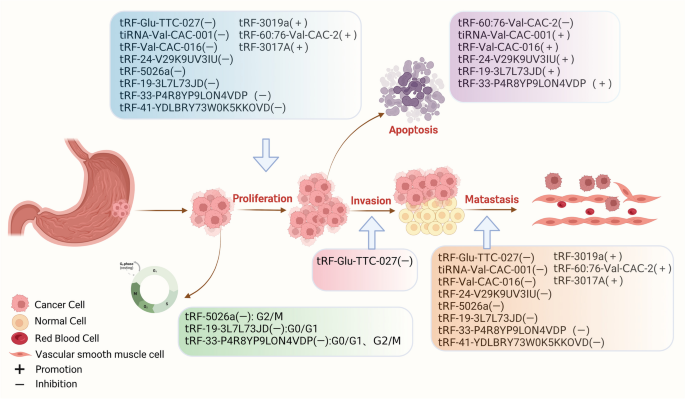
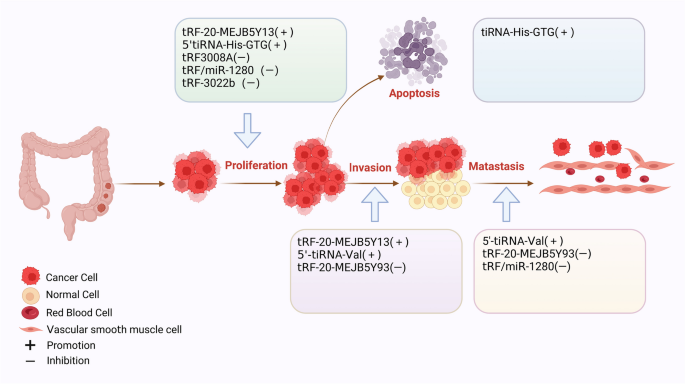
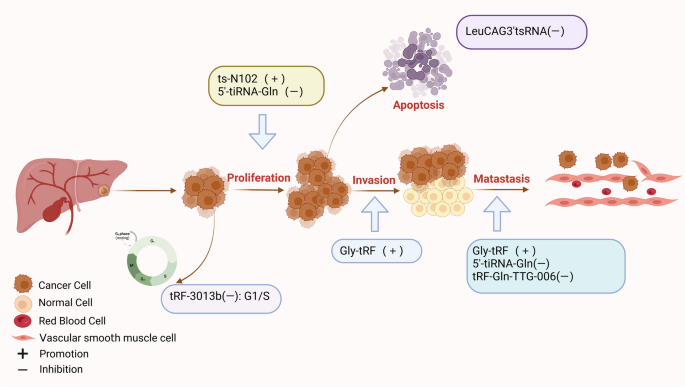
图书馆的src="http://www.rtryy.com/news/show/166171/ / / e.video-cdn.net/v2/embed.js " > 视频摘要 tRNA衍生的小非编码RNA (tsrna)是非编码RNA (ncRNA)家族的成员,可分为应激驱动RNA (tirna)和tRNA片段(trf)[1]。根据是否存在5'或3'序列,trf可以进一步分离为5'-tRF和3'-tRF,而tiRNA可以在5'-tiRNA和3'-tiRNA中分离[2]。根据酶切位点的不同,trf可被切割成多种亚型,如:tRF-5a、tRF-5b、tRF-5c、tRF-3a、tRF-3b、tRF-2等[3]。tsRNAs在不同的调控机制下发挥不同的生物学功能,影响人类疾病进展的不同阶段[4]。同时,在各种信号分子的调控下,tsRNAs参与遗传和表观遗传调控、RNA沉默、转录和逆转录调控、RNA稳定性调控和翻译调控[5,6,7,8]。早期研究已经证实tsRNAs存在于细菌、真核生物和古细菌中[9,10]。研究人员发现,由于测序技术的快速发展,tsRNAs普遍存在于酵母、布鲁氏锥虫、拟南芥和小鼠等多种物种中[11,12,13,14]。 近年来的研究表明,tsRNAs的生物学功能与多种生物过程密切相关,特别是在人类细胞中。人类的许多疾病(癌症和非癌症疾病)都是由于tsRNAs的失调而发生和发展的,如胃癌中的tRF-23-Q99P9P9NDD,结直肠癌中的tRF-26-P4R8YP9LOND,口腔纤维化中的trf - gly - ttc -016,急性胰腺炎中的tRF3-Thr-AGT[15,16,17,18]。消化系统疾病是临床上最常见的疾病之一,其患病率逐渐上升[19]。每年至少有10万患者首次接受胃肠道疾病治疗[20]。特别是消化系统癌症具有高发病率和高死亡率的特点[21]。消化系统是全球十大癌症中的四种,即肝细胞癌(HCC)、胃癌(GC)、食管癌(EC)、结直肠癌(CRC)。在十大癌症相关死亡原因中,有一半是消化系统肿瘤:CRC、HCC、GC、EC和胰腺癌(PC)[22]。 因此,消化系统癌症是全球癌症相关死亡的主要原因[23,24,25,26,27]。随着肿瘤标志物的检测和胃镜、结肠镜医学技术的普及,以及手术治疗、放化疗、靶向治疗、免疫治疗、综合治疗策略的应用,消化系统肿瘤的预后有了相对改善,但长期治疗效果仍不理想,严重威胁着人类的健康[28,29]。因此,提高肠癌的总体生存率需要在开发新的抗癌药物的过程中及早发现。最近,一些研究证实tsRNAs在各种消化系统肿瘤中具有重要的生物学功能,包括EC、GC、CRC、PC、口腔癌、下咽癌和肝胆系统肿瘤[30,31,32,33,34,35,36]。tsRNAs通常位于血浆/血清中。这在非侵入性癌症诊断中具有显著优势,因此tsRNAs有望作为早期检测癌症的生物标志物[15,34]。此外,本研究表明tsRNAs在评估癌症预后和作为新的治疗靶点方面具有重要的应用前景[37]。此外,tsRNAs在口腔纤维化、口腔菌群紊乱、肝损伤、脂肪肝、胰腺炎等良性消化系统疾病中也有重要的生物学贡献,具有良好的临床应用前景[17,18,38,39,40]。 本文就tsRNAs的起源、分类、调控机制及其在消化系统疾病中的生物学功能进行综述。我们集中研究了tsRNAs作为消化系统疾病诊断和预后分子标志物的临床潜力。 tsRNAs源于trna,是长度为18-40个核苷酸(nt)的短链ncRNAs,通过成熟trna或trna前体(pre- trna)的加工重组而成[7]。不同的tRNA来源和酶切位点允许tRNA形成一系列具有不同结构和作用的tsrna,这些tsrna属于trf和tirna[41]。其中,5′-tRF (14-30 nt)和3′-tRF (18-22 nt)分别随机通过5′端和3′端产生[42,43,44]。如图1所示,在tRNA成熟过程中,细胞核内的前tRNA的5'和3'端在RNase P和RNase Z的影响下被切除;相应地[45,46],RNase z特异性内切酶裂解催化3'端产生tRF-1 (14 - 30nt)[47]。D环上的切割位点包括tRF-5a (14-16 nt)、tRF-5b (22-24 nt)和tRF-5c (28-30 nt),以及T环上的切割位点包括tRF-3a和tRF-3b、5'-tRF和3'-tRF,反映了tRNA成熟结构产生5'-tRF的位置[48]。rna有31-40个核苷酸长,是通过切割反密码子环产生的。这些tirna通常分为5'-tiRNA和3'-tiRNA两半。i-tRF (18 - 50nt)和tRF-2是在T环和D环的两个位点同时裂解产生的[49]。 图1 tsrna的起源和分类。从5'和3'端衍生的trf称为5'-tRF和3'-tRF。在细胞核中的前trna中,RNase z特异性核内裂解催化3 '端产生tRF-1。在成熟的tRNA结构中,5'-tRF可以在D环上的切割位点被切割,包括tRF-5a、tRF-5b和tRF-5c。3’-tRF可以在T环的裂解位点产生,包括tRF-3a和tRF-3b, 5’-tRF和3’-3’-tRF。tirna是在反密码子环上切割形成的,可分为5'-tiRNA和3'-tiRNA。i-tRF和tRF-2是在T环和D环的两个位点同时切割形成的 近年来,随着生物信息学的普及和技术的进步,利用测序和基因组技术发现了许多独特的tsRNAs[50]。它们可以在精液、尿液、血清和血浆等体液中大量存在[34,51,52,53]。一些研究已经提供证据表明,失调的tsRNA表达谱与消化系统疾病的发生/进展有关[54]。本节我们将从癌症和消化系统温和疾病两个角度讨论tsrna相关研究的进展。 GC中的tsRNAs 全球肿瘤相关死亡的第三大流行驱动因素是预后不良的胃癌,尽管应用了各种传统和现代治疗方法,包括手术、靶向治疗、化疗和免疫治疗[55,56,57]。目前,tsRNAs作为新的GC诊断/预后生物标志物或治疗靶点的潜力正受到越来越多的研究。失调的tsRNAs可以通过多种机制功能促进或抑制GC的发生和进展。如图2所示,总结了tsRNAs在GC中的生物学功能。 图2 tsRNAs在GC中的生物功能。tsRNAs可影响胃癌细胞的增殖、侵袭、转移和凋亡 促进tsrna的生物功能 在许多方面,tsRNA可以促进GC的进展。GC组织中丰富的tRF-Val直接与真核翻译延伸因子1 α 1 (EEF1A1)蛋白结合,促进其易位至细胞核,改善EEF1A1与MDM2蛋白的相互作用,促进肿瘤蛋白53 (p53)泛素化,这些都有助于GC的进展[58]。除了rna结合蛋白(RBP)机制外,tsRNAs还可以通过与靶基因的结合发挥调控作用。在GC细胞和组织中,tRF-3019a高度过表达[59]。它可以与Argonaute 2 (AGO2)蛋白相互作用,靶向并调控F-box蛋白47 (FBXO47)基因表达,促进肿瘤细胞增殖/转移。此外,胃癌组织中tRF-3017A表达的显著增加可能通过阻断新的表皮生长因子样2 (NELL2)基因而增加胃癌恶性[60]。值得注意的是,tsRNAs促进GC的不同机制可能为临床应用提供了不同的前景。 抑制tsRNAs的生物功能 Shen等通过RNA测序发现,GC组织中tRF-33- p4r8yp9lon4vdp (tRF-33)明显降低。细胞功能实验表明,tRF-33能够抑制GC细胞的增殖、迁移和周期进程,并能促进细胞凋亡,提示tsRNA具有调节GC进程的潜力[61]。另一项研究发现,在GC组织中低表达的tRF-19-3L7L73JD可阻断G0/G1期细胞周期,抑制细胞增殖和迁移,抑制细胞恶性进展[62]。此外,研究发现过表达tRF-41(也称为tRF-41- ydlbry73w0k5kkovd)对肿瘤细胞的迁移和增殖具有抑制作用。此外,它还可以通过下调磷酸腺苷-硫酸磷酸合成酶2 (PAPSS2)来促进细胞凋亡[63]。值得注意的是,tsRNAs可以调节相关信号通路,影响GC的恶性进展。tRF-Glu-TTC-027分子在GC组织中明显下调,主要在细胞质中下调。tRF-Glu-TTC-027分子可抑制丝裂原活化蛋白激酶(MAPK)通路相关蛋白的表达,从而阻碍GC的进展[32]。另一项研究表明,tRF-5026a可通过PI3K/AKT(磷酸肌醇-3激酶)信号通路驱动G2/M细胞周期阻滞并阻碍GC进展[64]。zhang等发现tiRNA-Val-CAC-001通过调节Wnt/β-catenin信号通路抑制GC细胞的增殖和转移,从而阻碍GC的进展[65]。同样,Dong等人发现tRF-24-V29K9UV3IU通过抑制Wnt信号通路在GC进展中发挥抑癌作用[66,67]。根据研究,tRF-Val-CAC-016在GC组织中明显下调。这种下调可以有效地阻碍癌细胞的发展。进一步的研究揭示了tRF-Val-CAC-016如何通过结合钙电压门控通道亚单位α 1 D (CACNA1d)基因抑制MAPK通路,调控其表达[68]。综上所述,tsRNAs可通过多种信号通路调控GC细胞的增殖和转移,调控GC的恶性进展,为GC的诊断和治疗提供新的见解和靶点。 tsrna的诊断潜力 一些研究表明,tsRNAs在胃癌诊断中具有很好的潜力。胃癌患者血清中tRF-23-Q99P9P9NDD[15]和tRF-17-WS7K092[69]的表达水平明显高于胃炎患者和健康人。有趣的是,胃癌患者术后它们的表达明显降低。tRF-23-Q99P9P9NDD的曲线下面积(AUC)为0.783,诊断敏感性为63%,特异性为86%,高于碳水化合物抗原724 (CA724)、碳水化合物抗原199 (CA199)、癌胚抗原(CEA)等传统标志物的AUC。然而,当这四种标志物一起用于胃癌检测时,AUC更高,为0.862。此外,tRF-23-Q99P9P9NDD的AUC为0.724,对I/II期GC的敏感性为60%,特异性为85%,也优于CEA、CA199和CA724。合并AUC为0.819[15]。同样,经ROC分析,tRF-17-WS7K092对健康人鉴别GC的AUC为0.819,也高于CA724、CA199和CEA的AUC。四种标记组合后,AUC增加到0.882。另一个来自tRNA-Gln-TTG的位于6q24.2染色体上的tsRNA Hsa_tsr016141在胃癌的血清和癌组织中也出现上调,AUC值为0.692,略高于CEA(0.654)和CA199(0.621)[70]。这些结果表明,一些高表达的tsRNAs可能是GC的可靠诊断标志物。 相比之下,GC患者血浆中tRF-33- p4r8yp9lon4vdp (tRF-33)和tRF-27- 87r8wp9n1e5 (tRF-27)的表达水平明显较低。胃癌患者术后血浆中它们的水平明显升高[71,72]。tRF-33在准确区分早期GC与健康个体方面的AUC为0.757[71]。健康人晚期胃癌的AUC为0.766,而胃炎患者早期和晚期胃癌的AUC分别为0.717和0.635。GC患者组织和血浆中tiRNA-5034-GluTTC-2水平均明显降低,AUC分别为0.835和0.779。合并结果时,AUC为0.915[73]。此外,经ROC分析,血浆中tRF-19-3L7L73JD的AUC为0.6230 [62],GC组织中tRF-5026a的AUC为0.908[64]。另一项研究显示,tRF-3019a在区分健康组织与伴有I-II期、III-IV期、淋巴结转移及低病理分化的胃癌中的AUC分别为0.796、0.665、0.530和0.677[59]。这些研究表明,这些低表达的tsRNAs在GC诊断中具有有效的应用价值。对上调和下调的tsRNAs进行联合ROC分析将有可能获得一组理想的诊断标记。 tsrna的预后价值 越来越多的证据表明,tsRNAs在胃癌中具有良好的预后价值。GC患者血清中tRF-23-Q99P9P9NDD的表达与晚期T分期、TNM分期、淋巴结转移及神经血管侵犯密切相关。此外,Kaplan-Meier曲线显示,低水平tRF-23-Q99P9P9NDD的GC患者总生存时间更长[15]。另一项研究报道,GC患者血清中tRF-17-WS7K092表达水平与淋巴结状态、TNM分期、神经血管浸润有良好的相关性。生存分析表明,该tsRNA低表达的患者具有良好的生存结局[69]。在晚期胃癌患者中,tRF-33的下调已被证明与预后不良呈正相关,血清tRF-33的降低与较差的生存率相关[71]。高水平tRF-27的患者肿瘤较大,Ki67水平较高,生存期较短。此外,COX分析表明,tRF-27是胃癌患者的独立预测因子。胃癌组织中tRF-31- u5ykfn8dydzdd (tRF-31)显著上调,且与术前肿瘤大小呈正相关。与低水平的患者相比,高水平的tRF-31患者的生存率较低。COX多因素分析显示,tRF-31是一个独立的预测因子,显示其对GC死亡风险的强大预测价值[74]。tgf -5026a下调与胃癌患者肿瘤较大、TNM分期较晚、转移到其他淋巴结相关,COX回归分析认为其是独立的预后指标。此外,低水平组比高水平组的存活时间更长[64]。hsa_tsr016141与分化程度降低、T分期晚期、淋巴结转移阳性相关。hsa_tsr016141表达增高的胃癌患者预后较差[70]。这些发现表明它们可能对胃癌患者具有潜在的预后意义。 CRC内的tsrna 结直肠癌是所有癌症中死亡率第三高的癌症。新发病例和死亡人数显著增加[22]。越来越多的证据表明,一些tsrna在结直肠癌的发展中具有生物学意义。如图3所示,总结了tsrna在CRC中的生物学功能。 图3 tsrna在结直肠癌中的生物功能。tsRNAs可影响结直肠癌细胞的增殖、侵袭、转移和凋亡 促进tsrna的生物功能 有研究报道一些tsrna可能在结直肠癌中起致癌分子的作用。Xiong等通过RNA测序在结直肠癌中鉴定出16种异常的tsRNAs。功能富集表明,tRF-25-P940KK5Y93、tRF-24-NMEH623K25、tRF-30-XSXMSL73VL4Y、tRF-26-P4R8YP9LOND和tRF-29-P27JPJ60MVJY可能通过维生素代谢途径促进CRC的发展,而tRF-29-QU7BPN6ISBJO和tRF-27-Q99P9P9NH5N通过cGMP-PKG途径加速CRC的发展[16]。另一种tsRNA,名为tRF-20-MEJB5Y13,来源于Dicer1(一种核糖核酸酶),通过上皮-间质转化(EMT)途径促进结直肠癌的增殖和转移[33]。此外,tRNA-Val的一种衍生物5'-tiRNA-Val在结直肠癌组织中表达上调,可加速结直肠癌细胞的侵袭和转移,但不影响结直肠癌细胞的增殖[75]。5'-tiRNA-His-GTG长度为35 nt,主要存在于细胞质中。它在结直肠癌组织中显著增加,通过AGO的互补作用使大肿瘤抑制激酶2 (large tumor suppressor kinase 2, LATS2)沉默,进而通过抑制Hippo信号通路促进结直肠癌的进展[76]。对这些tsRNAs的深入研究可能为结直肠癌的治疗提供新的靶点。 tsRNAs的抑制生物功能 据报道,一些下调的tsrna在结直肠癌的发展中起致癌分子的作用。Luan等发现tRF-20-MEJB5Y93可通过靶向cludin -1,干扰EMT抑制结直肠癌转移[77]。此外,他们证实它可以结合4957-4976区并沉默MALAT-1的表达,MALAT-1可以通过与富含丝氨酸和精氨酸的剪接因子2 (SRSF2)相互作用调节1A染色体结构维持的选择性剪接(SMC1A),从而阻断结直肠癌转移[78]。tRF3008A是由3 ' -tRNAval衍生而来,可与Argonaute相关复合物结合,降低FOXK1的稳定性,导致FOXK1下调,阻断Wnt/β-catenin信号通路,抑制CRC进展[79]。tRF/miR-1280由tRNALeu和pre-microRNA产生,它可以结合JAG2的3'UTR区域并抑制其表达,进而调控Notch信号通路,最终减少CRC的转移和形成;此外,Notch信号通路的失活抑制了癌症干细胞(CSC)的表型,如Gata1/3和miR-200b的直接转录沉默[80]。Yang等报道,tiRNA-His-GTG可促进细胞凋亡增加,但不影响细胞活力[81]。tRF-3022b可以直接靶向LGALS1和巨噬细胞迁移抑制因子(MIF),从而抑制M2巨噬细胞极化和CRC进展[82]。从大肠杆菌中提取的tRNA-Val (UAC)和tRNA-Leu (CAG)具有细胞毒性,可显著抑制结直肠癌细胞的克隆生成能力[83,84]。这些发现揭示了一些tsRNAs的抑制生物功能,这些tsRNAs有望成为CRC的新的潜在治疗靶点。 tsrna的诊断价值 近年来,越来越多的tsRNAs在CRC样本中被发现异常表达,为这些患者提供了潜在的诊断价值。tRF-18-8R1546D2、tRF-22-WE8S68L528R1546D2、tRF-22-WB86Q3P92和tRF-22-WE8SPOX52已被Zhu等人发现在CRC组织中表达异常,并用于进一步建立新的诊断模型。ROC分析显示AUC值为0.93[85]。5'-tRF-GlyGCC是一种31nt片段,是人体体液中含量非常丰富的tsRNA,在结直肠癌患者血浆中显著上调[86]。5′-tRF-GlyGCC的AUC为0.882,显著高于CEA(0.762)和CA199 (0.557);联合诊断的AUC(0.926)最高[86]。tRF-VAL-TCA-002和trf -ph - gaa031在结直肠癌组织中表达明显上调,诊断AUC分别为0.7554和0.7313[87]。tRNA-Gly衍生的tRF-3022b在结直肠癌组织中过表达。血浆样本中分离CRC患者与健康受试者的tRF-3022b AUC为0.7684,组织样本中tRF-3022b AUC为0.8266,具有较强的微创诊断价值[82]。虽然这些tsRNAs在CRC的诊断中有各自优异的价值,但是通过对这些tsRNAs的联合分析,可以期待更好的诊断效果。 tsrna的预后价值 tsrna在结直肠癌的预后中显示出潜在的价值。Zhu等人构建了基于tRF-33-PSQP4PW3FJI0W、tRF-18-HSRVK7D2、tRF-18-H9Q867D2、tRF-33-PSQP4PW3FJIKW、tRF-16-I3FJQSD、tRF-32-O7M8LOMLQHWU3的CRC预测模型。高风险评分患者的生存时间明显较短。多因素分析证实,该预后模型在结直肠癌患者中是一个独立的指标[85]。5'-tiRNA-ProTGG下调,是结直肠癌的独立预后因素。5′-tiRNA-ProTGG阳性表达患者的无病生存期(DFS)明显短于阴性表达患者,肿瘤复发率明显增高,总生存期明显短于阴性表达患者[88]。tRF-phe-GAA031和tRF-VAL-TCA-002与总生存率呈显著负相关,证明它们可能是CRC中极好的预后生物标志物[87]。此外,CRC患者血清5′-tiRNA-Val水平明显高于健康对照组,其表达与肿瘤淋巴结转移呈正相关,可作为评估淋巴结转移状态的生物标志物[75]。总的来说,这些tsRNAs在CRC患者的预后中有很好的应用前景。 tsRNAs在肝胆癌中的作用 胆道肿瘤包括GBC、CCA和HCC[89]。虽然全身免疫治疗和靶向治疗提高了生存率,但晚期患者的预后并不理想[90]。因此,寻找新的有效的生物标志物对肝胆癌尤为重要。如图4所示,总结了tsRNAs在肝胆癌中的生物学功能。 图4 tsRNAs在肝胆肿瘤中的生物功能。tsRNAs可影响肝癌细胞的增殖、侵袭、转移和凋亡 促进tsrna的生物功能 与胃癌和结直肠癌不同,tsrna在肝癌中的作用尚不清楚。然而,一些研究报道tsRNA在促进肝细胞癌的进展中起重要作用。长度为22 nt的leucag3’tsrna在小鼠胆囊癌模型中高表达,可调节RPS28的翻译,减少细胞凋亡[91]。Gly-tRF在肝胆肿瘤细胞中表达上调,促进肝肿瘤干细胞的EMT。具体来说,Gly-tRF可以直接靶向Nedd4家族相互作用蛋白2 (NDFIP2),这是AKT信号通路的一个组成部分,导致迁移和EMT[92]。另有研究发现,5′trf - gly可结合并沉默癌胚抗原相关细胞粘附分子1 (CEACAM1)的表达,进而促进肝癌细胞的增殖、迁移和侵袭[93]。分析显示,ts-N102可能抑制hs-mir-215的表达,并在肝癌中作为原癌基因。功能富集分析表明,ts-N102可能参与脂肪酸的合成和代谢[94]。 tsRNAs的抑制生物功能 生物体就像一个高度复杂的机器,tsRNAs的抑制生物功能也存在,以维持生命的内稳态。由tRNAGln - TTG产生的5'-tiRNA-Gln在肝癌中过度表达。它可以与真核翻译起始因子(EIF4A1)形成g -四重结构后相互作用,从而抑制肝癌细胞的增殖和迁移[95]。tRF-3013b被下调并抑制GBC的进展。它直接作用于肿瘤蛋白P63 regulated 1 like (TPRG1L),下调NF-κB、c-myc、CDK2,使GBC细胞停留在G1/S期[96]。tRF-Gln-TTG-006已被证明具有肿瘤抑制作用,但其机制尚不清楚[97]。 此外,一些促进或抑制作用尚未明确的tsRNAs在肝胆癌中也起着至关重要的作用。如Li等人在CCA中发现了许多差异表达的tsRNAs,并选择了具有统计学意义的tRF-34-JJ6RRNLIK898HR、tRF-38-0668K87SERM492V和tRF-39-0668K87SERM492E2进行进一步评价。这些tsRNAs均来源于tRFGluTTC,可靶向ruppel样因子家族16 (KIF16)调节脂肪酸形成并影响CCA进展[98]。 在肝癌中异常表达的tsRNAs也具有潜在的临床应用价值。患者血浆中tRNA-ValTAC-3、tRNAGlyTCC-5、tRNA-ValAAC-5、tRNA-GluCTC-5的表达明显高于健康对照组,提示其在肝胆癌中的微创诊断价值[34]。HCC患者血清中tRF-Gln-TTG-006水平明显升高。对来自不同医院的两组HCC患者与健康对照进行ROC分析,AUC分别为0.919和0.875。合并两个队列时,AUC为0.875。这些结果表明HCC患者中tRF-Gln-TTG-006水平存在高度的个体异质性。此外,进一步评价发现tRF-Gln-TTG-006的AUC为0.858,在分化I期优于AFP,提示其是HCC的有效生物标志物[97]。5'tRF-Gly在肝胆癌组织和细胞中表达上调,与肿瘤生长、远处转移、分化不良呈正相关。表达较多5′trf - gly的个体生存时间较短[93]。ts-N102在肝癌组织中表达上调,ts-N7、ts-N94、ts-N84、ts-N37表达下调。为了区分原发肿瘤和健康肿瘤,ROC分析显示,基于这5种tsrna的诊断效果的AUC值为0.88。此外,采用ts-N20、ts-N21、ts-N22、ts-N36、ts-N37、ts-N44、ts-N64 7种tsRNAs构建风险模型,证明该模型能准确预测肝胆癌预后[94]。这些发现提示,这些上调或下调的tsRNAs可能在肝癌的诊断和预后中具有潜在的应用价值。 其他癌症中的tsrna tsRNAs还参与其他消化系统癌症的发生和进展,包括口腔癌、下咽癌、食道癌和PC癌。如图5所示,总结了tsRNAs在这些癌症中的生物学功能。 图5 tsrna在其他癌症中的生物功能。tsRNAs可影响OSCC、EC和PC细胞的增殖、侵袭、转移和凋亡 tiRNA-Val-CAC-002是5'-tRF家族的成员,位于1号染色体上。它在口腔鳞状细胞癌(OSCC)组织中弱表达,并可能通过PI3K/AKT通路促进OSCC的发展[17]。tRF-21- vby9pykhd (tRF-21)衍生自tRNAGlyGCC,是一种PC肿瘤抑制因子。tRF-21的减少促进akt2 /1介导的异质核核糖核蛋白L (hnRNP L)磷酸化,进而磷酸化hnRNP L增强与hnRNP L- dead -box解旋酶17 (DDX17)的相互作用,形成选择性剪接复合体,促进PC细胞恶性。此外,用tRF-21模拟物治疗可以抑制PC小鼠异种移植物的肿瘤生长和转移,这表明tRF-21可能作为抑制PC进展的治疗靶点[99]。tRF-Leu-AAG可以直接靶向并抑制上移码蛋白(UPF1)的表达。敲低tRF-Leu-AAG可抑制PC细胞的增殖、转移和侵袭[100]。tRF-Leu-CAG-002和tRF-Pro-AGG-004可促进PC细胞迁移和肿瘤生长[101]。既往研究表明,TRG-GCC的5'端形成的tRNA-GlyGCC-5表达增加,可促进EC的增殖和转移[31]。 tRNA86ArgTCT衍生的tRF-20-S998LO9D在口腔舌鳞癌(SCCOT)中显著上调。高水平tRF-20-S998LO9D个体生存时间较短,预后AUC值为0.669[102]。 tRF-1:30-Lys-CTT-1-M2在下咽癌中高表达,与肿瘤分期晚、分化差相关。ROC分析显示,其AUC为0.9172,具有较好的诊断价值。此外,COX多因素分析发现它是肺转移的独特危险因素[30]。tRNA-GlyGCC-5鉴别EC患者与健康个体的AUC为0.878,表明该tsRNA具有潜在的诊断作用[31]。 PC患者血液中tRF-Leu-CAG-002和tRF-Pro-AGG-004显著升高。与健康个体鉴别PC患者的AUC值分别为0.9和0.78。在鉴别I/II期患者时,CEA和CA19-9联合诊断的疗效明显优于CA19-9。这些tsRNAs水平高的患者生存率较低,可作为术后肿瘤监测的标志物[101]。tsRNA-MetCAT-37和tsRNA-Val-TAC-41在PC患者的组织和血清中明显升高。联合CA199的AUC值分别为0.949和0.947,对PC与健康人群的鉴别具有较好的诊断价值。此外,tsRNA-ValTAC-41的上调与肿瘤晚期和远处转移呈正相关[103]。在PC组织中,tRF-Pro-CGG表达下调,其AUC为0.92。tRF-Pro-CGG低表达与淋巴结转移阳性和肿瘤分期晚期相关。Kaplan-Meier分析显示,低水平tRF-Pro-CGG的PC患者总生存时间较短,提示其预后和诊断价值[35]。此外,AS-tDR-000064、AS-tDR-000069、AS-tDR-000102和AS-tDR-001391在PC中存在差异表达。然而,这些临床应用价值需要进一步的研究来证实[104]。 综上所述,tsRNA与特定的基因和蛋白相互作用,在消化系统癌症中显示出致癌和抑制功能(图6)。我们的综述可能为tsRNA在消化系统癌症中的功能和临床应用前景提供更好的视角。 图6 tsRNAs在癌细胞中的调控机制。tsRNAs与蛋白质和基因相互作用,在消化系统癌症中显示生物学功能 tsRNAs在良性消化系统疾病中也有关键作用。本节将讨论tsRNAs与口腔粘膜下纤维化(OSF)、口腔菌群紊乱、肠易激综合征(IBS)、良性肝脏疾病和急性胰腺炎(AP)之间的关系(表1)。 表1前任良性消化系统疾病中tsRNAs的表达、临床价值及功能 口腔良性疾病 OSF是一种癌前病变,其中1.5-15%可成为口腔恶性肿瘤[112]。因此,开发和应用分子生物标志物治疗和预防OSF尤为重要。研究发现,tRF-Gly-TCC-016在OSF组织中表达上调,常靶向IL-13和TGF-β家族细胞因子,通过细胞因子-细胞因子受体相互作用和cAMP信号传导影响OSF的形成;tRF-Gly-TCC-016被认为是OSF的分子标记物和治疗靶点[17]。相反,tiRNA-Val-CAC-002在组织中下调。tiRNA-Val-CAC-002的靶基因主要富集于膜和细胞器中,其主要作用围绕ATP代谢和对β淀粉样蛋白生物反应的调节[17]。 核梭杆菌(Fusobacterium nucleatum, Fn)在口腔疾病的治疗中至关重要[38]。他等人发现一些tsRNAs可以靶向感染,抑制Fn中的核糖体靶向,并调节微生物-宿主环境[113,114]。他们发现tsRNA-000794和tsRNA-020498可以通过阻止氨基酸合成蛋白质来限制Fn的生长[113]。更有趣的是,感染Fn的NOKSI(正常口腔角质形成细胞)细胞在特定时间点产生更多的tsRNA-000794和tsRNA-020498[113]。这表明口腔角质形成细胞和口腔细菌之间存在潜在的相互作用,这些相互作用是由特定的tsRNAs介导的。然而,本研究并未探索tsRNAs与细菌相互作用的机制,而这对于研究口腔疾病与微生物群的关系至关重要。 肠易激综合症 肠易激综合征伴腹泻(IBS-D)是一种功能性肠道疾病[115],但其发病机制尚不清楚[116]。研究表明,tiRNA-His-GTG-001、tRF-Ser-GCT-113和tRF-Gln-TTG-035与功能性肠道疾病有关[105]。tiRNA-His-GTG-001位于第9染色体tRNA-His基因上。tiRNA-His-GTG-001的长度为34 nt,来源于tRNA-His-GTG的5'端,在IBS患者中高表达。生物学功能分析显示,tiRNA-His-GTG-001靶基因与谷氨酸代谢、gaba能突触、TNF-α信号通路、胰岛素抵抗等关键通路和代谢过程具有显著相关性。分析表明,tiRNA-His-GTG-001可靶向GABAB受体2 (GABBR2), GABBR2是一种主要的伤害感受分子,在疼痛调节中起重要作用[117]。因此,tiRNA-His-GTG-001可能通过靶向GABABR2的3'UTR调节肠易激综合征的易激性腹痛。tRF-Ser-GCT-113全长31nt,在肠易激患者肠黏膜内低表达,位于M染色体MT-TS2基因上。tRF-Ser-GCT-113与肠易激的某些症状有关,如腹胀和腹泻。5-羟色胺2C受体(HTR2C)和SAM和SH3结构域蛋白1 (SASH1)是tRF-Ser-GCT-113的靶标基因,与应激敏感性和临床情绪障碍相关[118],提示tRF-Ser-GCT-113可能通过作用于HTR2C和SASH1引发患者出现腹部不适、腹胀等症状。而tRF-Gln-TTG-035的长度为14 nt,在IBS患者肠黏膜中低表达。与tRF-Gln-TTG-035相关的靶基因与PPAR信号通路相关。它可以结合并作用于toll样受体4 (TLR4)和γ -氨基丁酸受体相关蛋白(GABARAP),抑制它们的翻译。进一步研究表明,tRF-Gln-TTG-035通过调节TLR4和GABARAP影响脑-肠神经元免疫介导的信号传导,导致腹部扩张程度增加。对患者组和正常对照组进行ROC分析,发现tirna - hs - gtg -001、tRF-Ser-GCT-113、tRF-Gln-TTG-035的AUC值分别为0.921、0.792、0.889,提示其有可能成为有效的诊断标志物和新的治疗靶点[105]。 良性肝病 肝损伤和衰竭是常见的危及生命的疾病,需要多学科合作治疗[39,119]。 tRF-Gln-CTG-026属于tRF-1,它与TSR1(预rRNA加工蛋白TSR1同源物)相互作用,抑制TSR1和18S rRNA的作用,从而减少核糖体组装,减少整体蛋白合成,减轻肝损伤,这表明它可能是一个治疗靶点,可以为治疗肝损伤提供一种新的途径[107]。 到目前为止,HBV(乙型肝炎病毒)的流行率仍然很高[120],全世界估计有3.5亿人患有慢性HBV感染,而超过1.6亿人患有慢性丙型肝炎病毒(HCV)。不幸的是,在此期间,超过100万患者死于与慢性病毒性肝炎相关的并发症[121]。长度为30-35个核苷酸的5' trhs (5'-tiRNA)在这些病毒感染过程中起着至关重要的作用[106]。研究人员发现,5'tRHVal和5'tRHGly在HBV和HCV感染的肝组织中表达显著升高。此外,他们的研究小组发现5'tRHVal和5'tRHGly不参与肝癌细胞中蛋白质合成的控制。5'tRHs在病毒性肝炎中的作用值得进一步研究[106]。 在HBV相关急性、慢性肝衰竭(ACLF)中,tsRNA-20和tsRNA-46的表达下调[108]。生物学功能分析表明,它们与MAPK信号通路、乙型肝炎通路以及其他与代谢、增殖和凋亡相关的通路具有显著的相关性。构建了包括tsRNA-20和tsRNA-46以及其他ncrna在内的mrr - rna特征来检测ACLF。这种mrr - rna标记可以熟练地区分HBV-ACLF病例和正常对照。研究人员将这些人分为训练组和验证组。在训练队列中,该特征显示出诊断效率,AUC为0.75,灵敏度为86.25%。在验证队列中,mrr - rna标记有效地将HBV-ACLF病例与正常受试者区分开来,AUC为0.787,灵敏度为74.29%。结果提示tsRNA-20和tsRNA-46可能成为疾病诊断的重要指标[108]。 脂肪肝是一种代谢相关疾病;无有效药物预防[122]。在酒精性脂肪性肝病(AFLD)的研究中,一项测序分析显示,gly - trf全长29-34 nt,在其tRNA前体的5 '端反密码子环上发生了切割。它们在AFLD患者(长期摄入乙醇的小鼠)中表现出显著的上调。进一步研究发现补体C3可以通过调节细胞色素P450酶CYP2E1的表达促进Gly-tRFs的产生。Gly-tRF参与sirtuin 1 (SIRT1)信号通路调节脂质代谢,Gly-tRF抑制剂将成为AFLD的治疗策略[40]。 在一项非酒精性脂肪性肝病(NAFLD)的研究中,tRF-47- 58zzjqjyswryvmmv5bo (tRF-47)的表达显著上调。进一步研究发现,tRF-47表达增加可增强自噬,减少促死亡信号,减少脂质沉积,表明其可通过促进自噬过程来达到治疗NAFLD的效果[109]。研究表明,tRF-3001b在NAFLD患者中表达明显升高。进一步的实验分析表明,tRF-3001b通过Prkaa1途径抑制自噬,导致脂质形成,加重NAFLD的进展。在NAFLD小鼠模型中,tRF-3001b升高,可抑制自噬基因Prkaa1的表达,加重NAFLD[110]。因此,沉默tRF-3001b可减少肝脂质形成,从而改善NAFLD的发展,这可能是治疗NAFLD的新途径。Huang等人观察到NAFLD患者的肝组织和血浆中一些tRNA片段显著增加[111]。其中,tRF-Ala-CGC-006、tiRNA-His-GTG-001和tRF-Val-CAC-005因其高表达水平而被选择进行全面研究。生物学功能分析显示它们主要参与脂质代谢的控制。用于区分NAFLD和非NAFLD样本的AUC值分别为0.875、0.840和0.868。它们具有良好的诊断价值,可作为生物标志物。值得注意的是,有研究证实,tRFGluTTC等trf可抑制脂肪生成,并伴有aP2、PPARγ等脂肪生成转录因子的下调,提示其在脂质代谢中的重要作用[123]。 美联社 AP是一种越来越常见的慢性炎症[124,125]。在AP动物模型中,我们发现tRF3-Thr-AGT表达下调。当tRF3-Thr-AGT表达升高时,AR42J细胞中的几种炎症因子(如caspase1、IL-1β和IL-18)被抑制,这表明它可能参与胰腺炎期间炎症的启动。生物信息学分析表明,ZBP1、CD44和BTG2是tRF3-Thr-AGT的下游靶点。它能够附着在ZBP1的3'UTR上,通过z - dna结合蛋白1 (ZBP1)影响细胞热裂解和炎症的发生,并控制ZBP1/NLRP3通路,防止细胞热裂解和炎症。这些数据表明,tRF3-Thr-AGT对胰腺炎的进展具有抑制作用,是AP的保护因子[18]。在该团队的另一项研究中,他们建立了胰腺腺泡细胞内胰蛋白酶原激活模型(PAITA),发现tRF3-Thr-AGT是该模型中的枢纽trf。其表达水平明显降低。tRF3-Thr-AGT也与胰蛋白酶原激活有关,为诊断和治疗AP提供了一种新的生物标志物[126]。 摘要。 介绍 tsrna的起源和分类 tsRNAs在消化系统疾病中的作用 有限公司 结论与展望 数据和材料的可用性 缩写 参考文献。 致谢。 作者信息 道德声明 # # # # # ccDownload: /内容/ pdf / 10.1186 / s12964 - 023 - 01341 - 8. - pdf