动物以各种方式选择和与环境互动,包括确保它们的生理机能处于最佳状态,有可能获得猎物,以及可以避开捕食者。选择的平衡往往是相互矛盾的,这取决于个人的生活史和状况。普通的蜥蜴(胎生蜥蜴)有产卵和生育的血统,并显示出各种各样的背部图案和颜色。颜色和繁殖模式如何影响景观上的栖息地选择决策尚不清楚。在这项研究中,我们首先测试了共同发生的雄性和雌性胎生和卵生普通蜥蜴在其微栖息地选择上是否存在差异。其次,我们测试了一只蜥蜴的背部颜色是否与它所遇到的微栖息地内的日光浴地点选择相匹配,这可能与伪装和隐蔽有关。
研究发现,蜥蜴对生境的利用不同于其他可获得的生境,这表明蜥蜴积极选择其微生境的组成和结构。与雄性相比,雌性出现在树木较多、裸露地面较少的地区;我们推测,这可能是为了更好地伪装,减少怀孕期间被捕食的风险,因为怀孕期间雌性的流动性较差。微生境的使用也因胎次模式而异:胎生蜥蜴在开花植物密度更高的地区被发现,而胎生蜥蜴在更潮湿、苔藓更多的地区被发现。这可能与胎生动物和卵生动物对产卵地点的不同栖息地偏好有关。我们发现一个个体的背部颜色与其晒地的基质颜色相匹配。这可能表明个体可能会选择他们的日光浴地点来优化微栖息地的伪装。此外,所有的个体都在靠近掩蔽物的地方晒太阳,我们认为这可以用来躲避捕食者。
我们的研究表明,普通蜥蜴可能会主动选择它们的微栖息地和晒地,平衡生理需求、逃跑反应和伪装作为一种躲避捕食者的策略。这因胎次模式、性别和背部颜色而异,表明个体优化策略受到种群内个体间变异的影响,并由与生活史相关的进化差异决定。
在他们的环境中,任何动物物种的个体都有很多选择来度过他们的时间。这些微生境选择决策主要取决于寻找食物、参与社会互动和躲避捕食者之间的权衡[1,2,3,4]。其中,捕食者躲避可能起着最重要的作用,因为不谨慎的选择可能导致立即死亡[3]。为了避免被捕食,个体通常面临两种选择:躲藏或逃跑。伪装,即动物的颜色和图案与其周围环境之间的相似性,是适应的一个显著例子,已成为进化研究中的模型系统[5,6,7,8,9]。对野生动物伪装的研究表明,生活在各种颜色微栖息地的猎物要么在不同的微栖息地之间妥协,要么专门生活在一个微栖息地以优化捕食者的躲避[7,10]。例如,在树皮上休息的飞蛾,如胡椒蛾(Biston betularia),对与自己颜色相似的微栖息地表现出偏好[11,12],甚至会重新定位身体以增强伪装[13,14,15]。伪装也可能取决于个体所遇到的不同微栖息地的多样性。例如,沙漠蜥蜴(Sceloporus magister)在它们使用的两个或多个微栖息地之间折衷了一种颜色[7]。
在鳞片中,背部的图案和颜色主要是为了与周围的底物相匹配而进化的[16,17,18]。然而,颜色和图案也可能进化为对性信号和环境压力的反应,包括温度和对紫外线辐射的保护[19,20,21,22,23,24]。例如,在Montivipera raddei毒蛇复合体中,背部图案颜色的变化与热因素有关,包括温度和太阳辐射[25]。然而,在个体的一生中,捕食者的躲避策略可能会发生变化,这取决于个体发生的颜色、冲刺速度(逃跑表现)的变化,或者由于繁殖和获得配偶而导致的行为变化[26,27,28]。总体而言,研究发现颜色、图案、性别和动物个性之间存在复杂的关系,这表明在这种变异的进化过程中存在相互依赖的权衡[23,29,30]。
提供伪装的机会是栖息地选择的重要考虑因素,但微栖息地也至关重要地提供了免受捕食者的保护[31,32],遇到猎物的机会[4,33],或对变温动物的体温调节益处[2,34]。例如,植被覆盖和较短的躲避距离使动物能够避免被发现,降低捕食风险[32,35]。另一方面,露天的日光浴场所为变温动物提供了最大限度地暴露在阳光下的最佳机会,因此对温度调节有好处[36,37],但更容易暴露于潜在的捕食者[38]。因此,个体需要仔细选择他们在不同类型的微栖息地中度过的时间。此外,这些选择可能取决于个体的性别和季节。例如,雌性在怀孕期间通常活动较少,因此需要随着飞行速度的降低而调整它们的行为[39,40]。
虽然已经有充分的证据表明,蜥蜴选择具有特定特征的微栖息地(例如,为了体温调节[41,42,43,44]或捕食者躲避[1,45]),但很少有研究测试同一物种内表现出种内外观差异的个体是否会主动选择微栖息地内的特定背景。最近的研究确实表明,在某些物种中,个体可能会主动选择与自己颜色相匹配的栖息地。例如,在地面筑巢的鸟类中,个体将自己的颜色和蛋与它们筑巢的栖息地相匹配[46],当它们自己与背景(而不是它们的窝)的匹配不佳时,它们更容易逃跑[47]。居住在希腊岛屿的爱琴海壁蜥(Podarcis erhardii)个体将自己的颜色与背景相匹配,雌性和捕食风险较高的地区的效果更强[15]。因此,个人对场地使用的决策可能涉及当地环境的可用性和生物的颜色变化。
普通蜥蜴(活蜥蜴)是一种小型的、生活在地面上的蜥蜴,分布在欧亚大陆。除了在陆地爬行动物中拥有最大的分布范围外,它在生活史的生殖方面也表现出惊人的差异:一些种群是产卵的(卵生),而另一些是活产的(胎生)[48,49]。普通蜥蜴的繁殖季节从早春开始到夏季结束,在此期间,怀孕的雌性体重可能会增加一倍[50]。这一时期也是普通蜥蜴最容易受到捕食者攻击的时期,这些捕食者通常依赖于视觉线索,比如猛禽、蛇和一些哺乳动物。
普通蜥蜴在个体间的颜色和背部图案上表现出相当大的差异[51],在浅棕色的背景上有网状或线形图案,或介于网状和线形图案之间的中间图案[23]。个体的背部图案在第一年形成,然后在其一生中保持稳定[23]。普通蜥蜴没有主动的颜色变化来匹配或伪装特定的基质,如变色龙等其他蜥蜴[52]。身体状况和杂合性等特征在普通蜥蜴的择偶中起着重要作用[53,54],但背部的颜色或图案似乎不是关键的线索。相反,网状和线性形式在种群中出现的频率取决于环境条件和基因遗传[23]。然而,无论是雄性还是雌性,背部颜色和图案在景观上的关系都没有得到详细的研究。
在本研究中,我们评估了生殖季节合生的卵生和胎生蜥蜴的微生境选择。我们首先测试了普通蜥蜴的微栖息地选择是否在它们所处的环境中是特定的。此外,我们检查了他们的选择是否与胎次方式(卵生或胎生)和性别有关。最后,我们在考虑大小和性别差异的情况下,测试了普通蜥蜴背部的颜色和图案是否与晒太阳部位的颜色有关。我们测试的假设是:(i)普通蜥蜴偏爱微栖息地的特定特征,(ii)卵生和胎生的普通蜥蜴在微栖息地选择上存在差异,(iii)怀孕的雌性会遇到与雄性不同的微栖息地,(iv)背部颜色与蜥蜴晒日光浴地点的颜色相匹配,(v)背部图案与晒日光浴地点的颜色有关。
普通蜥蜴显然在取样研究区域内选择了特定的微栖息地。该地区可用的微栖息地类别的相对比例与蜥蜴使用的微栖息地之间存在显着差异(图1和图2)。这一点可以通过首选简单蜥蜴模型(其中假设普通蜥蜴是一个同质群体,表1,第2行和第3行)的DIC(偏差信息标准)估计至少比零模型低5来证明。假设在所有9种生境类型中,有普通蜥蜴的地点与随机地点之间没有差异(表1,第1行)。特别是,普通蜥蜴往往出现在被子植物、凋落叶、苔藓、水和木材比例较高的地区,而覆盖物、岩石、草和光秃秃地的比例较低的地区(表1;附加文件1(表S1);图2).如果选择一个简单的模型来区分蜥蜴和随机可用的地点,那么普通蜥蜴会出现在草(和)比例较低、凋落叶(和)比例较高的地区。当岩石存在时,岩石在不同种群(和)之间的比例相似,但在蜥蜴(和)附近更可能完全没有。
图1
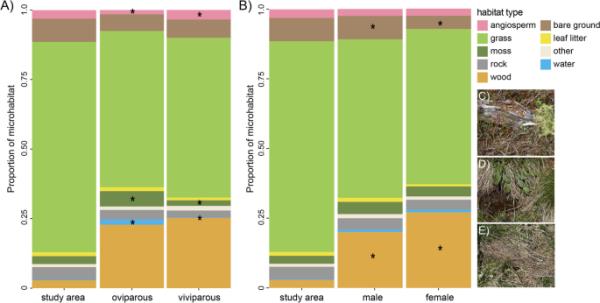
(A)卵生和胎生蜥蜴以及(B)雄性和雌性蜥蜴所选择的微栖息地中(“研究区”)总体上遇到的栖息地类型的相对比例。宇称模式(A)或性别(B)不同的生境类型用星号标记。图C - E)显示了不同栖息地的例子,包括常见蜥蜴的典型晒地,如C)石南丛中的干原木,D)水边的干草丛,E)草地上的干草地
图2
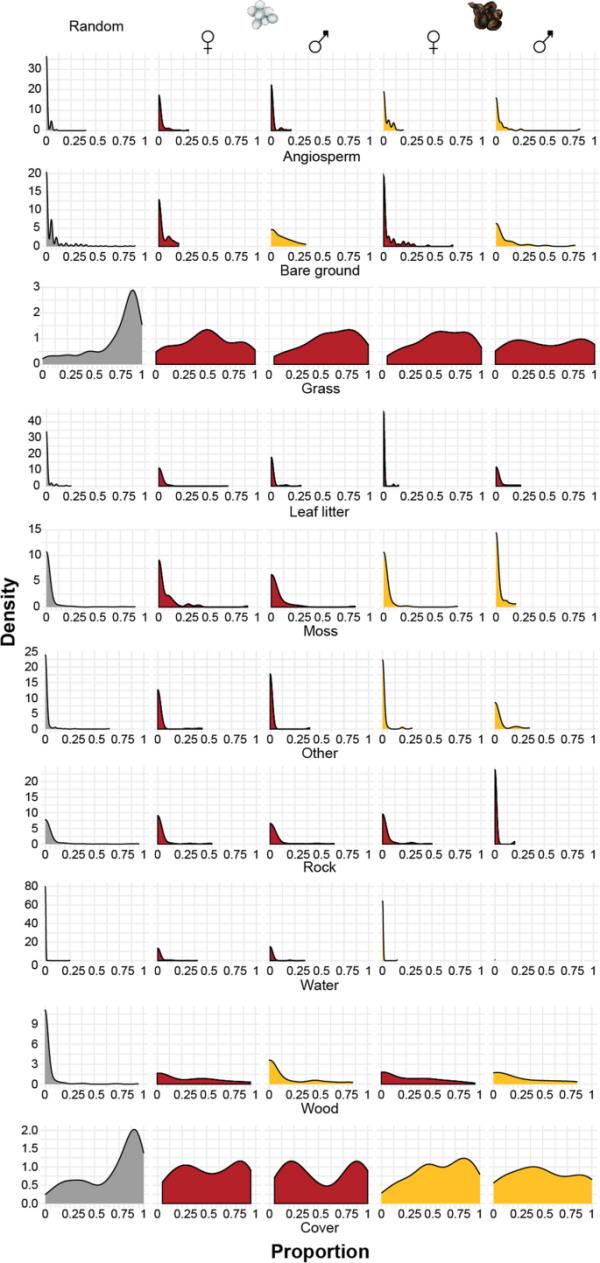
在解释各生境成分差异方面表现最佳的生境成分和因子的比例(根据DIC评估)。核密度图中的不同颜色表示在最佳表现模型中显著不同的因素。蜥蜴所使用的栖息地的所有组成部分都与研究区域不同(“随机”;灰色的颜色)。红色和黄色编码的组合表明,宇称模式或性别是另一个重要因素。例如,在胎次模式是一个重要因素的情况下,第二列和第三列(卵生雄性和雌性;红色编码)不同于第四和第五列(胎生雄性和雌性;编码的黄色)
表1零和一膨胀beta (ZOIB)回归模型的偏差信息准则(DIC)解释了蜥蜴附近和随机选择的微栖息地类型比例的变化泰德样方
对于九种栖息地类型中的六种,基于蜥蜴之间差异的更复杂的模型更受欢迎(表1)。与雌性和随机可用性(和)相比,雄性蜥蜴更有可能在附近()观察到没有裸露地面(比例=0)(附加文件1(表S1))。当存在时,两种胎次模式的雌性蜥蜴附近的木材比例都高于雄性蜥蜴(或随机蜥蜴)。与卵生蜥蜴相比,胎生蜥蜴更常与被子植物()联系在一起。在没有水的情况下,卵生蜥蜴比胎生蜥蜴或随机存在的蜥蜴更不容易被发现,当有水时,水在卵生蜥蜴周围环境中所占的比例更高(而和)。在没有苔藓的情况下,卵生的普通蜥蜴也比胎生的蜥蜴或随机存在的蜥蜴更不容易被发现。最后,胎生的蜥蜴更有可能完全有或没有覆盖物(比没有覆盖物的比例高1),同时,卵生的普通蜥蜴对栖息地中可用覆盖物的偏好表现出双峰分布,而胎生的蜥蜴则表现出单峰分布(图2)。
背侧颜色与胎次模式、性别、体长和体重有关(表2)。更具体地说,胎生和体型较大的个体往往具有较高的颜色(表2;附加文件2(表S2);附加文件3(图S1))。此外,胎生的、雄性的、较大的和重量较轻的蜥蜴往往有更强烈的颜色(模型1(饱和度);表2;附加文件2(表S2))。最后,与胎生个体相比,胎生个体的背部颜色较浅(估计=-3.12,t1,250=-3.34, P < 0.001;表2;附加文件2(表S2))。
表2蜥蜴背部颜色、图案、微生境和晒地颜色之间的关系。所示为所有显示与反应显著相关的统计模型分析了无变量。缩写:Num.=分子;窝。=分母;Df=自由度
背侧图案与体长和性别有关(模型2;表2;图3;附加文件2(表S2))。其中,雄性呈网状分布的比例高于雌性(估计值=1.068,z1387=3.05, P < 0.01;表2;图3B)和较小的蜥蜴更可能具有线性模式(估计=0.071,z1,387=2.79, P < 0.01;附加文件2(表S2))。具有网状图案的个体通常具有较浅的背部颜色(模型3;F2,299=3.28, r2=0.015, p < 0.05;图3 c;附加文件2(表S2))。
图3
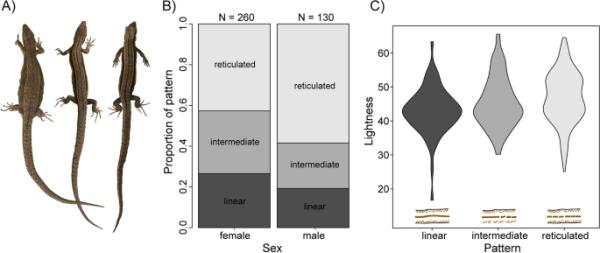
背部图案因性别而异,并与背部颜色有关。A三个雌性的网状,中间和线性模式的例子(从左到右)。B网状、中间或线状背纹的个体比例,分别显示雄性和雌性。C网状个体在背部颜色上的总体亮度得分较低
普通蜥蜴的背色范围为27至51(平均36.8),晒点范围为20至69(平均47.3),栖息地范围为33至69(平均54.9)。生境与晒地色相呈正相关(模型4;估算值=0.39,t1160=8.75, R2=0.331, P < 0.001;),并且晒地的变量多于栖息地(标准差:10.6 vs. 7.2)。普通蜥蜴的体重范围为25.1至64.9,晒地范围为12.9至79.4,总体微生境范围为17.3至47.6(附加文件4(图S2))。晒地亮度与生境总体亮度呈显著相关(模型5;估计=0.33,t1,160=10.81, R2=0.427, P < 0.001;表2;附加文件2(表S2)),但分布有所不同,日光浴地点比栖息地更轻(平均值:36.8比30.1),亮度变化更大(标准差:12.7比6.6;附加文件4(图S2))。
背部颜色与蜥蜴选择的晒日光浴微栖息地的颜色相关(模型6)。具体而言,蜥蜴背部区域的色调与栖息地的色调相关(估计=0.21,t1162=5.29, R2=0.151, P < 0.001;表2;图4)和晒地(估计值=0.09,t1162=3.00, R2=0.05, P < 0.01;表2;因此,蜥蜴通常倾向于选择与自己颜色相匹配的栖息地和日光浴地点。此外,蜥蜴倾向于选择比自身颜色稍深的地方晒日光浴(估计=-0.10,t1162=-2.14, R2=0.03, P < 0.05;表2;图4)与颜色相反,背部图案,即线性或网状,与晒地或栖息地颜色无关(N=161, P > 0.1;附加文件2(表S2))。
图4
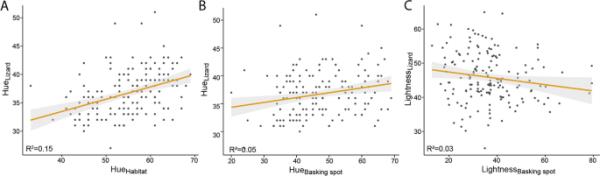
普通蜥蜴背部的颜色与其栖息地的颜色呈正相关关系。图中显示了蜥蜴的颜色与(A)栖息地和(B)日光浴地点的颜色的对比。图(C)说明了蜥蜴的亮度与其周围晒日光浴地点的亮度之间的关系。效应量包括在线性模型的R2中
摘要
背景
结果
讨论
结论
材料与方法
数据和材料的可用性
参考文献
致谢
作者信息
道德声明
补充信息
搜索
导航
#####
我们的研究结果表明,在自然环境中,普通蜥蜴积极选择它们的微栖息地,可能是为了优化体温调节和生理需求,同时最大限度地降低风险。蜥蜴选择所有栖息地成分的比例明显不同于它们在邻近地区的可用比例。我们发现,普通蜥蜴通常会避开裸露的岩石和裸露的地面。通过这种方式,我们认为它们可以避免在这样暴露的栖息地被发现和被捕食的风险,这是一个重要的考虑因素,因为它们是相当缓慢的地栖蜥蜴。普通蜥蜴对环境的湿度要求相对较高[55,56],这反映在它们对有更多苔藓的潮湿栖息地的偏好上。此外,凋落叶和被子植物是首选,可能提供了逃避的选择,而不被捕食者发现。
作为晒日光浴的地方,普通蜥蜴经常使用木材,相对于邻近地区,它们的微栖息地也丰富了木材。木材通常与蜥蜴的背景颜色很好地匹配,并且还提供了快速进入逃生避难所的裂缝[27]。此外,木材升温很快,因此为温度调节提供了最佳选择[57]。与环境中随机分布的植被相比,植被覆盖较少的微栖息地也更受欢迎,大概是因为普通蜥蜴需要晒太阳的地方,而较高的植被减少了可用于体温调节的太阳辐射量。
除了这些普遍的偏好外,小生境的选择还取决于个体的生活史:雄性和雌性、卵生和胎生蜥蜴的选址不同。雌性喜欢有良好的日光浴选择(更多的树林)和较少的开放空间(“裸露的地面”)的栖息地。在繁殖季节,雌性会怀孕并降低速度[40,58,59],因此可能需要相对“安全”的微栖息地,在那里它们可以迅速达到最佳体温,但也可以在短距离内逃到一个藏身之处。雌性普通蜥蜴待在避难所附近,表现出较低的飞行容忍度[27],可以抵消它们在怀孕期间速度下降的影响,以避免被捕食。通过在微生境选择实验之外进行运行性能测试,可以在未来使用更大更平衡的样本量来测试这种潜在的性别偏见。
奇偶模式也与普通蜥蜴的微生境选择有关。具体来说,产卵的普通蜥蜴是在苔藓比例较高、开花植物比例较低的潮湿微生境中发现的。这证实了先前的一项研究,即产卵的普通蜥蜴生活在更潮湿的栖息地[60]。我们认为其原因可能是为了获得最佳发育,卵窝需要潮湿的环境。在苔藓下和潮湿的枯木中发现了产卵的普通蜥蜴的卵窝(个人观察),在人工饲养的情况下,如果有选择,卵窝通常存放在湿润的苔藓下[50]。在微生境中发现胎生蜥蜴,被子植物的比例较大,而其他成分在胎次模式之间没有差异。总之,我们表明个体的性别和生活史(雄性与雌性,卵生与胎生)与普通蜥蜴在自然环境中的微栖息地选择有关,可能是为了匹配生理需求和与捕食风险的权衡。这表明,长期的进化差异在个体的行为策略中发挥作用。
我们的研究结果进一步表明,与背景基质相比,普通蜥蜴选择特定的微栖息地和日光浴地点,这与它们的背部颜色有关。相对于其他选择,个体倾向于选择更接近自身颜色(以色相衡量)的栖息地和晒地颜色。这一结果与其他研究一致,表明蜥蜴将背景与自身的颜色相匹配,以减少捕食者的发现[15,61,62]。然而,就鸟类捕食者而言,我们的结果应该谨慎解释,因为我们没有测量蜥蜴的全部光谱和它的背景。蜥蜴和鸟类在它们的视觉光谱中都覆盖了紫外线,我们在这里没有测量。与蜥蜴的颜色与其栖息地之间的正相关相反,蜥蜴晒日光浴的地方的亮度往往比它自己的颜色要暗。一种可能的解释是,蜥蜴在晒太阳时优化了它们的热环境。由于较深的表面有利于更有效的体温调节,与较深的个体相比,较浅的个体可能需要更多的时间来升温[63],因此在晒日光浴时选择较深的表面可以抵消这种差异。总之,我们的研究结果表明,蜥蜴个体面临着加热效率和匹配背景之间的权衡。
普通蜥蜴是如何根据自己的肤色选择栖息地和日光浴地点的,这是一个有趣的问题。蜥蜴有可能通过卷曲来直观地评估自己背部的颜色,这是它们在晒太阳时通常采取的一种行为。另一种说法是,背部颜色的基因座可能与对最适合其颜色和图案的栖息地的行为偏好有关。另一种可能是,蜥蜴从以前与捕食者的接触中学会了优化它们的伪装行为[64]。晒日光浴地点在颜色、基质和植被组成上与其他即时微生境不同,这强烈表明它们相对于即时可用的微生境是被主动选择的。
我们的发现也证明了蜥蜴在个体之间的颜色差异。特别是不同胎次模式、性别、体长和体重个体的颜色强度差异显著。卵生个体与胎生个体在所有三个背色变量上都存在差异:胎生个体的色调和饱和度值较大,而亮度值较低。虽然这些差异是微妙的,但它证实了现场的印象,即卵生个体往往比胎生个体略轻。此外,在体型较大的个体中,色调向更大的值移动,可见从深棕色到浅棕色的变化。这与普通蜥蜴在个体发育过程中经历颜色变化是一致的,而与体重和整体健康的相关性值得进一步研究。网状图案的个体倾向于背部颜色较浅。我们还发现图案与性别有很大的关系,网状图案在男性中更常见,而线性图案在女性中更常见。另一项研究表明,在壁虎(Podarcis muralis)种群[24]和不同的常见蜥蜴谱系[23]中,雌性的线性图案比例更高,性别之间的差异与此一致。先前的研究比较了普通蜥蜴的背部模式和探索行为,发现线性个体在没有覆盖的栖息地斑块之间的过渡似乎更大胆,并且与网状个体相比具有更高的迁移率[23,29]。因此,有可能复杂的行为特征与背部模式有关,尽管在这里我们没有发现这与不同的晒太阳地点选择有关。
由于背侧图案对微生境选择没有显著影响,我们认为相对较暗的线性形态在雌性中更为丰富可能是一种温度调节适应。在晒日光浴期间,较暗的线性图案可能使腹侧柱更快升温,并改善体温调节[23,24]。雌性出现线性模式的频率较高,这可能是由于在怀孕期间,体温调节所产生的能量增加,有助于逃脱捕食者[24,65,66]。一般来说,图案的纹理和几何形状,连同身体的颜色,可以代表那些背景视觉特征的随机样本,并有助于在晒太阳时伪装[61,67]。图案也可能通过打破身体线条或合并身体边缘以与基底匹配而造成破坏性伪装[61,68,69,70]。虽然在这个相对较小的空间尺度上,我们没有发现普通蜥蜴的背部图案和晒日光浴地点之间的联系,但图案可能与更大尺度上的栖息地有关,例如当栖息地彼此之间存在较大差异时(例如,高山草甸与泥炭地草甸相比)。
总的来说,我们的主要发现表明,普通蜥蜴有选择性地选择它们的微栖息地,这相对于立即可用的地点来说是非随机的,可能是为了平衡生活史依赖的生理需求、伪装和捕食者躲避。普通蜥蜴优先出现在增强匹配的背景中,这可能是它们在自然微栖息地抵御捕食者的伪装,同时保持在掩蔽物附近,以便快速逃脱。进一步的工作需要了解蜥蜴在这些位点选择中的主动选择,以及伪装的背景匹配是否有利于生存。其他研究表明,基因控制和视觉输入可能是适应性背景选择的基础,个体可以学会区分伪装和非伪装背景[15]。虽然这项研究测试了真实的个体颜色(通过相机RGB值捕获),并且不依赖于人类视觉系统所指示的颜色,但其他研究依赖于以人为导向的指数[71,72,73]。基于人类视觉系统的隐性测量可能有偏差,或者与其他动物的测量不同,因为不同的动物群体的视觉系统差异很大[74,75,76,77]。最后,使用比数码摄影更广泛的光谱反射数据,并将背景和猎物的颜色直接与潜在捕食者的视觉系统联系起来,将有利于爬行动物隐影的未来研究[15,61,78,79,80]。
这项研究是在奥地利加里恩阿尔卑斯山脉的盖勒河谷进行的。研究区包括6个被可见屏障(如河流、道路、森林)分隔的地点,总面积约0.3 km2,海拔范围为1350 ~ 1550 m。该地区在夏季(6 - 9月)用于放牧牛羊,栖息地以高山草甸为主。该研究区是产卵和生育的普通蜥蜴之间为数不多的接触区之一,这是两种遗传分化和不同的系统发育谱系[48,49,60]。
2016年5月至7月,共捕获了394只个体,并通过雄性存在阴茎隆起而雌性没有阴茎隆起来进行性别鉴定。个体在标准化条件下拍摄,以准确量化颜色信息(见下一节)。使用数字卡尺(Moore和Wright)测量鼻部到排气口的长度(SVL)和尾部长度(TL),使用Pesola微线弹簧秤测量重量。雌性蜥蜴饲养在单独的28 × 19 × 14 cm的塑料terraria中,提供藏身之处、湿苔藓、水和食物(粉虫、蟋蟀),并可随意取食[50]。在产卵或分娩后,从每只雌性身上取下一根尾巴夹进行基因分析,然后在捕获时将其释放。活产的雏鸟和它们的母亲一起被放生,而孵出的卵则在24°C下孵化,并在它们的母亲孵化后被捕获时释放。通过表型(即,如果个体产下一窝卵或产下活的幼崽)和SNP基因分型来评估胎次模式[49]。构建ddRADSeq文库,并在Illumina HiSeq 4000机器上以2 × 150bp的读取长度对其进行测序。使用STACKS对个体进行解复用[81],并将其映射到常见蜥蜴参考基因组[82]。对snp进行筛选和提取,要求个体间存在率> 50%,最小等位基因频率为10%。使用K=2对所有个体进行admix分析,以解释该地区遇到的卵生和胎生系统发育谱系。如果隶属度值(Q)在0.1和0.9之间,则将个体记录为混合并排除。较高或较低的值分别记录为纯卵生或胎生。
在标准设置下(固定ISO: 160,光圈:8,曝光补偿:0),使用佳能70D和60mm固定镜头,以RAW格式拍摄图像,以保留色彩信息。每张图像包括一个“X-rite ColorChecker”板,用于标准化图像之间的颜色并捕获准确的颜色[83,84]。首先,制作每个图像的数字负片(DNG)副本,然后通过颜色检查软件(X-rite ColorChecker)运行,该软件以配置文件的形式建立准确的颜色基础。接下来,在分析之前,将该配置文件应用于原始图像,以确保标准和准确的颜色结果。使用Adobe Photoshop CC 2015处理图像。将轮廓文件应用于图像后,为X-rite颜色棋盘上的off-white正方形选择白平衡,随后将所选正方形值的红绿蓝(RGB)线性设置为240纳米(nm)[85]。最后,记录背部条纹周围区域的RGB值,其中颜色采样区域包括从前腿到后腿的背部;侧翼被排除在外。计算色相、饱和度和明度(HSL)值来评估颜色。色相是主色调的近似值(从红色到蓝色),而饱和度测量其强度,明度通过量化添加到色相中的黑色或白色的数量来反映颜色的暗或浅。我们的解释主要集中在色调(H)和亮度(L)上。我们注意到,虽然我们的方法允许在人类视觉光谱(300-700 nm)内无偏提取颜色,但它忽略了紫外线光谱内的颜色。因此,这些颜色分布对哺乳动物捕食者具有代表性,但蜥蜴(包括活体动物[86])和鸟类[75,87]的光谱灵敏度范围并未完全覆盖。
从图像中,背部条纹被眼睛分类为线性,网状或中间的身体前部和后部。由于之前曾报道过蜥蜴身体前后部分的差异[24],因此分别记录了这两个区域的背侧图案。对394只个体(264只雌性和130只雄性)的背纹进行了分类,并对背色进行了量化。
栖息地测量是在每个个体被发现晒太阳并被捕获的同一地区进行的。晒太阳的地点被定义为蜥蜴第一次被发现和静止的地方。在用佳能70D和60毫米镜头捕捉蜥蜴后,立即从上面拍摄了栖息地的图像。每张图像包括一个0.5 × 0.5 m的样方,样方中间放置了大约0.15 × 0.15 m的日光浴位置,样方外面有一个X-rite ColorChecker板。栖息地图像在Adobe Photoshop中处理,使用与蜥蜴背部图像相同的颜色采样方法,并使用X-rite ColorChecker软件准备轮廓。记录样方中心0.15 × 0.15 m晒地的RGB值。在所有取样个体(N=394)中,共有165只(120只雌性和45只雄性)被取样进行背部着色和晒地着色。
生境被归类为由样方内包含草、苔藓、凋落叶、被子植物、木材、岩石、裸土、水或“其他”基质的区域组成。使用图像和覆盖在计算机上的栖息地图像上的网格(25个3厘米x 3厘米的网格正方形)推断栖息地类别比例。生境比例定义为每一类的方格数除以样方内的方格总数。将蜥蜴可以立即逃脱捕食的潜在掩护的比例量化为和样方中包括高草、木头和其他样方内个体可以快速躲藏的区域的比例。在394个个体中,有283个个体(185个雌性个体和98个雄性个体)对微生境的利用进行了分析。
此外,在整个研究区随机拍摄了240张生境图像,与晒地的基质进行了比较。研究区域可以划分为六个地理区域,通过划分道路、河流或森林等特征进行分隔[50],并在每个地理区域中确定随机地点,方法如下:从六个采样点的每个中心开始,通过向空中投掷一根棍子随机选择方向。手杖有一个指定的尖端,着陆后,手杖的尖端决定了行走的方向。经过20步后到达随机采样点,生成一个邻近但独立的微生境测量值。然后将样方与研究人员的脚垂直放置,并以与日光浴地点相同的方式拍摄图像。通过重复上一步确定下一个随机采样点,现在从当前随机采样的点开始。这整个过程在六个采样点中的每个点重复了十个随机采样点。现场小组的每个成员(N=4)执行此步骤一次,因此每个采样点包含40张随机栖息地测量图像(4人x 10个随机点)。整个研究区域共获得240张图像(6个采样点x 40张图像)。由于曝光过度,一张图像随后被排除。栖息地分类是通过图像量化的,方法与发现蜥蜴在晒日光浴的小栖息地相同。
为了评估普通蜥蜴的微栖息地选择,我们使用贝叶斯0 - 1膨胀β (ZOIB)回归对蜥蜴附近各种栖息地类型的存在、缺失或比例进行了建模[88]。ZOIB分布允许对包含0和1的比例数据进行建模。在这种情况下,响应变量被建模为伯努利分布和beta分布的混合,从中分别产生真正的0和1,以及0和1之间的值。概率密度函数为
Where and and。混合参数决定了伯努利分量或β分量占主导地位的程度,值越高表示0或1的数量越多。参数是从伯努利分量中得出的概率。对于没有观察到零或1的响应变量,分别固定为要么或。在我们的数据中出现了没有观测的生境类别,在这些情况下,混合参数决定了为零的概率,而不是使用beta分量建模的比例。连续的数据点,使用一个beta分布进行建模,参数化均值和精度。
针对每种生境类型,采用9种版本的ZOIB回归模型进行评估。第一个零模型包含一组参数,并适用于所有蜥蜴和随机地点。下一个最简单的模型允许在蜥蜴和随机地点之间变化(“简单蜥蜴模型”)。进一步的模型也允许在雌性和雄性蜥蜴之间,在卵生和胎生之间变化,最后在性别和胎次模式之间变化。除了null之外,每个模型都使用所有观测值共享的单个参数来实现,对于蜥蜴和随机站点则使用两个参数来实现。通过比较偏差信息标准(deviance information criterion, DIC)来进行模型比较[89],最佳模型被认为是DIC最低的模型,除非发现一个更简单的模型,其DIC分数相差小于2,在这种情况下,更简单的模型被优先考虑。
贝叶斯模型对每个观察类都使用最小信息先验来实现,其中类可以在简单模型中表示随机或常见的蜥蜴位点,或者在更复杂的模型中表示具有特定性别或奇偶模式的常见蜥蜴位点。对于每个观测类,,先验分布为:混合参数;对于逻辑回归中的概率;为回归均值;和回归精度。模型在JAGS语言中指定,并使用由R (R Core Team 2013)发起的JAGS v4.3.0[90]运行,使用包runjags计算DIC。缺口[91]。
接下来,我们评估了普通蜥蜴的背部颜色是否依赖于个体的任何属性(例如,奇偶模式、性别、长度、体重),以及它是否与它们的背景颜色匹配。我们从背色、晒地和栖息地的图像中提取HSL(色调、饱和度、亮度)值。接下来,使用广义线性模型(GLM)来确定平均背部颜色(以HSL测量)是否与个体的胎次模式、性别、体长和体重相关(模型1)。同样,使用二项分布的GLM来确定背部图案(网纹、中间或线性)是否取决于胎次模式、性别、体长和体重(模型2)。使用方差分析,我们测试了背部颜色是否与背部图案相关(模型3)。我们检查了整个微栖息地的颜色是否与晒地颜色相关(模型4)。然后,我们测试了平均背部颜色是否与晒地和性别相关(模型5),以及与整个微栖息地的颜色和性别相关(模型6)。因此,我们也测试了背部图案的颜色(模型7和8)。
DIC青睐的0 - 1膨胀β回归模型参数。
在蜥蜴的颜色和图案上进行的统计模型。显示了模型中任何显著相关变量的影响。
背部颜色在宇称模式之间是不同的,并且与体重有关。胎生(A)和体型较大(B)的个体比胎生和体型较小的个体具有更大的色相值。
A)更宽生境亮度直方图和B)晒地亮度直方图。
下载原文档:https://link.springer.com/content/pdf/10.1186/s12862-023-02158-2.pdf